Scene 1 (0s)
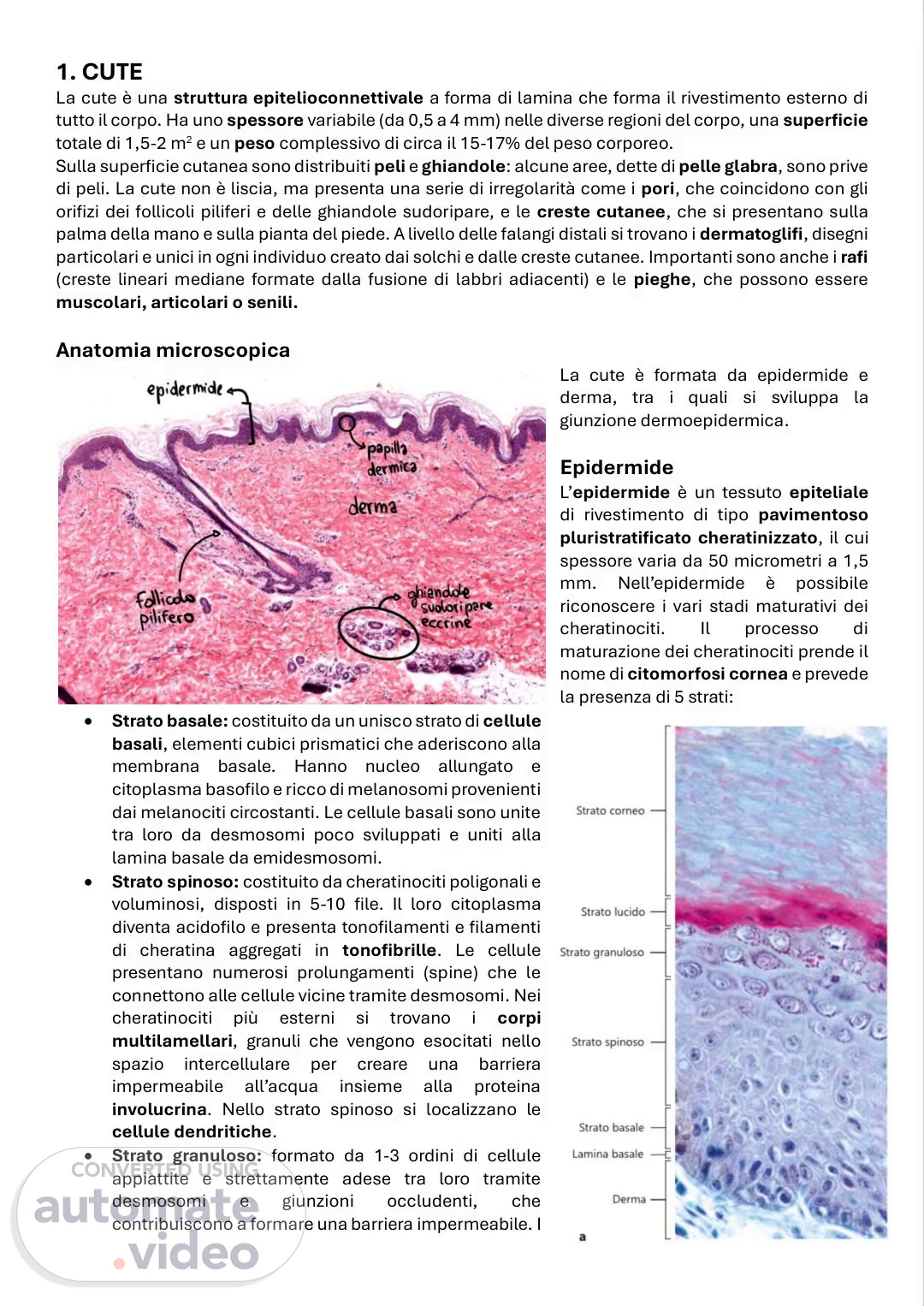
[Audio] 1. CUTE La cute è una struttura epitelioconnettivale a forma di lamina che forma il rivestimento esterno di tutto il corpo. Ha uno spessore variabile (da 0,5 a 4 mm) nelle diverse regioni del corpo, una superficie totale di 1,5-2 m2 e un peso complessivo di circa il 15-17% del peso corporeo. Sulla superficie cutanea sono distribuiti peli e ghiandole: alcune aree, dette di pelle glabra, sono prive di peli. La cute non è liscia, ma presenta una serie di irregolarità come i pori, che coincidono con gli orifizi dei follicoli piliferi e delle ghiandole sudoripare, e le creste cutanee, che si presentano sulla palma della mano e sulla pianta del piede. A livello delle falangi distali si trovano i dermatoglifi, disegni particolari e unici in ogni individuo creato dai solchi e dalle creste cutanee. Importanti sono anche i rafi (creste lineari mediane formate dalla fusione di labbri adiacenti) e le pieghe, che possono essere muscolari, articolari o senili. Anatomia microscopica La cute è formata da epidermide e derma, tra i quali si sviluppa la giunzione dermoepidermica. Epidermide L'epidermide è un tessuto epiteliale di rivestimento di tipo pavimentoso pluristratificato cheratinizzato, il cui spessore varia da 50 micrometri a 1,5 mm. Nell'epidermide è possibile riconoscere i vari stadi maturativi dei cheratinociti. Il processo di maturazione dei cheratinociti prende il nome di citomorfosi cornea e prevede la presenza di 5 strati: Strato basale: costituito da un unisco strato di cellule basali, elementi cubici prismatici che aderiscono alla membrana basale. Hanno nucleo allungato e citoplasma basofilo e ricco di melanosomi provenienti dai melanociti circostanti. Le cellule basali sono unite tra loro da desmosomi poco sviluppati e uniti alla lamina basale da emidesmosomi. Strato spinoso: costituito da cheratinociti poligonali e voluminosi, disposti in 5-10 file. Il loro citoplasma diventa acidofilo e presenta tonofilamenti e filamenti di cheratina aggregati in tonofibrille. Le cellule presentano numerosi prolungamenti (spine) che le connettono alle cellule vicine tramite desmosomi. Nei cheratinociti più esterni si trovano i corpi multilamellari, granuli che vengono esocitati nello spazio intercellulare per creare una barriera impermeabile all'acqua insieme alla proteina involucrina. Nello strato spinoso si localizzano le cellule dendritiche. Strato granuloso: formato da 1-3 ordini di cellule appiattite e strettamente adese tra loro tramite desmosomi e giunzioni occludenti, che contribuiscono a formare una barriera impermeabile. I.
Scene 2 (3m 8s)
[Audio] nuclei sono alterati o assenti e il citoplasma presenta i granuli cheratoialini, granuli privi di membrana rappresentati da filaggrina, proteina che aggrega i filamenti di cheratina in macrofibrille. Nello strato granuloso viene espressa anche la loricrina, proteina che crea ponti tra alcune proteine fissandole al rivestimento dei corneociti. Strato lucido: presente solo nelle regioni palmari e plantari, formato da 1-3 strati di cellule morte che appaiono traslucide nei preparati a fresco. La translucenza di questo strato è dovuta all'eleidina, sostanza ricca di lipidi e proteine che deriva dalla maturazione della cheratoialina. Il suo ruolo è quello di proteggere l'epidermide dall'azione di soluzioni acquose. Strato corneo: è lo strato più superficiale dell'epidermide. Risulta formato da corneociti con contorni irregolari, membrana ispessita e nucleo assente. Si rileva uno strato corneo disgiunto, in cui i desmosomi sono alterati, e uno strato corneo compatto, in cui i desmosomi sono in numero minore. Il citoplasma dei corneociti si disidrata e si riempie di filamenti di cheratina, la cui aggregazione è favorita da filaggrina e ponti disolfuro. Sulla superficie dei corneociti involucrina e loricrina permettono l'organizzazione del materiale esocitato dai granuli cheratoialini, determinando l'ispessimento della membrana plasmatica. Altri tipi cellulari che si rilevano nell'epidermide sono: Melanociti: cellule deputate alla sintesi dei pigmenti di melanina, responsabili della colorazione della cute. Sono situati nello strato basale e il loro numero corrisponde a un quarto di quello delle cellule basali. Sono dotati di prolungamenti che si estendono nello strato spinoso e presentano nel citoplasma granuli di pigmento scuro (melanosomi). Cellule dendritiche: rappresentano la prima linea delle difese cutanee, essendo cellule estremamente efficienti nel processare e presentare antigeni ai linfociti. Sono situati nello strato spinoso e presentano numerosi prolungamenti. Sono mobili e presentano le caratteristiche vescicole lageniformi striate, a forma di racchette da tennis e delimitate da membrana. Cellule di Merkel: hanno funzione di meccanocettori. Situate al confine tra epidermide e derma, hanno forma tondeggiante e citoplasma omogeneo e ricco di vescicole contenenti mediatori chimici. Si ritrovano raggruppate in formazioni dette menischi tattili. Giunzione dermoepidermica Il derma aderisce all'epidermide tramite la giunzione dermoepidermica. Si identifica con la membrana basale e ha un aspetto ondulato, dato dalla presenza delle papille dermiche. Presenta tre zone: Lamina densa: spessore di 20-50 nm, formata da una rete tridimensionale di fibrille collagene di tipo IV e laminina 5, che conferisce resistenza e flessibilità. Presenti anche fibronectina e proteoglicani. Lamina lucida: attraversata da filamenti di ancoraggio che assicurano le cellule epiteliali alla lamina densa. Contiene laminine ed altre glicoproteine necessarie all'adesione cellulare. Lamina fibroreticolare: presenta fibre collagene di tipo VII. Derma Il derma è una membrana biancastra formata da tessuto connettivo denso a fasci intrecciati con spessore variabile da 0,3 a 4 mm. La componente cellulare è formata prevalentemente da fibroblasti, mentre la componente amorfa è formata da acqua, elettroliti, glicoproteine e proteoglicani. Sono importanti in particolar modo l'acido ialuronico, che trattiene grandi quantità di acqua, e i proteoglicani solforati (dermatansolfato, eparansolfato e condroitinsolfato) a cui si deve la compattezza e la densità del tessuto connettivo. Il derma provvede al trofismo e al sostegno dell'epidermide, e in esso si sviluppano la maggior parte degli annessi cutanei. È divisibile in due strati: Strato papillare: presenta le papille dermiche, sporgenze coniche che aumentano la.
Scene 3 (7m 31s)
[Audio] Strato reticolare: costituito da fasci di fibre collagene e numerose fibre elastiche. Presenta dei fasci connettivali profondi, che delimitano spazi in cui si sviluppano follicoli piliferi, glomeruli di ghiandole sudoripare e lobuli adiposi. Peli Nel pelo si distinguono tre parti principali: Fusto del pelo: sottile bastoncello a sezione circolare o ellittica, può essere rettilineo o ondulato e ha un diametro variabile tra 20 e 500 micrometri. Presenta una superficie irregolare data dalla presenza di scaglie cornee embricate. Dall'esterno verso l'interno è composto da 3 sezioni: cuticola, corteccia e midolla. Follicolo pilifero e radice del pelo: il follicolo pilifero è una formazione sacciforme dilatata in profondità (bulbo del pelo) e più ristretta in superficie (istmo del follicolo pilifero), verso l'orifizio esterno sulla cute. A questo livello si ha lo sbocco di una ghiandola sebacea. La parte di pelo contenuta nel follicolo pilifero è detta radice del pelo e forma, insieme al follicolo e alla ghiandola sebacea annessa, il complesso pilosebaceo. La parete del follicolo pilifero è composta da strati epiteliali sovrapposti, mentre la sua porzione profonda è formata da: o Guaina esterna della radice, formata da un solo strato di cheratinociti a livello del bulbo e da più strati salendo verso la superficie. A livello dell'inserzione del muscolo erettore del pelo presenta il bulge del pelo, che accoglie un pool di cellule staminali o Guaina interna della radice, presente solo nella parte profonda del follicolo e formata da tre strati che terminano a livello dell'istmo, dove desquama e forma uno spazio che separa la cuticola del pelo dalla guaina interna. All'esterno della guaina esterna si trova la membrana vitrea, formato da due strati: uno profondo, formato da fibre collagene e fibre elastiche, e uno più prossimo all'epidermide formato da fasci di fibre collagene. Annessi del pelo Al follicolo pilifero sono generalmente annessi: Ghiandole sebacee: piccole, formate da uno o due alveoli, con il dotto escretore che sbocca a livello dell'istmo del follicolo pilifero; Muscolo erettore del pelo: origina dal tessuto connettivo papillare del derma e si inserisce sulla membrana vitrea del follicolo pilifero. La sua contrazione permette lo svuotamento delle ghiandole sebacee e l'orripilazione. Ghiandole sebacee Sono ghiandole acinose tubulo-alveolari contenute nello strato papillare del derma: si ha un numero di alveoli variabile da 2 a 20, che si aprono in un dotto escretore che si apre verso l'epidermide. Gli alveoli sono riccamente vascolarizzati e hanno una parete formata da una fila di cellule basali periferiche, cellule staminali che rimpiazzano gli elementi perduti. Il dotto escretore è formato da cellule epiteliali appiattite. Le ghiandole sebacee hanno una secrezione di tipo olocrino: le cellule si riempiono di gocciole lipidiche e vanno incontro a disfacimento, entrando a far parte del secreto, detto sebo. Il sebo ha pH leggermente acido ed è formato da materiale lipidico, detriti cellulari e batteri. Ghiandole sudoripare eccrine Sono ghiandole tubulari semplici di tipo glomerulare. La parte secernente è posta in profondità e forma un gomitolo (glomerulo), mentre il dotto escretore si apre sull'epidermide in corrispondenza di una cresta cutanea. Il glomerulo è composto da cellule mioepiteliali che rivestono uno strato di cellule epiteliali secernenti che si affacciano nel lume. Il dotto escretore invece è tappezzato da due strati di cellule epiteliali. Le cellule dello strato profondo sono simili.
Scene 4 (11m 33s)
[Audio] elettroliti. Nelle ghiandole sudoripare eccrine la produzione di secreto avviene senza distruzione cellulare. Il secreto è un liquido incolore formato principalmente da acqua e soluti vari, e la sudorazione è comandata da stimoli termici o emozionali. Il sudore è importante dal punto di vista della termoregolazione e della regolazione dell'equilibrio idrico dell'organismo. Ghiandole sudoripare apocrine Sono ghiandole tubuloglomerulari, costituite quindi da un glomerulo e un dotto escretore. Il glomerulo è posto nello strato reticolare del derma, il dotto escretore terminale è ampio e si apre spesso in un follicolo pilifero. Il tubulo è formato da cellule cubiche o cilindriche (in base alla fase secretoria) in rapporto con cellule mioepiteliali che poggiano su una membrana basale. Il citoplasma delle cellule in fase attiva presenta numerosi granuli di secreto, che si spostano verso la porzione apicale per essere emessi nel lume ghiandolare. Il sudore apocrino è un liquido acquoso, leggermente opaco e alcalino (pH tra 7 e 8). 2. MAMMELLA La mammella è un rilievo cutaneo pari e simmetrico posto sulla superficie anteriore del torace, ai lati della linea mediana. È situata lungo la linea del latte, linea che si porta dall'ascella al tubercolo pubico, lungo la quale si possono osservare piccole ghiandole soprannumerarie o eterotopiche. Tra le due mammelle si interpone un solco detto seno. La mammella contiene una parte ghiandolare, situata al di sopra della fascia di rivestimento del muscolo grande pettorale. È rivestita da cute molto sottile che presenta all'apice l'areola, zona rotondeggiante pigmentata al centro della quale sorge il capezzolo. Anatomia microscopica La principale ghiandola della mammella è la ghiandola mammaria. Ha forma discoidale e presenta un corpo costituito da 15-20 lobi, immersi in tessuto adiposo e separati da lamine connettivali. Ogni lobo si divide a sua volta in lobuli che contengono gli alveoli, unità secernenti. Ciascun lobo rappresenta una ghiandola tubulo-acinosa composta e fa capo a un dotto galattoforo, che all'interno del lobo si divide nei dotti extralobulari che raggiungono i lobuli continuando nei dotti intralobulari. I dotti intralobulari proseguono nei dotti alveolari e alla fine si immettono in gruppi di alveoli terminali. In superficie i dotti galattofori si dilatano nei seni galattofori, che fungono da serbatoi di riserva del latte. Nella donna non gravida in età riproduttiva si rileva un sistema di dotti ramificati, inglobato in una massa di tessuto fibroso interlobulare denso e alla periferia si rileva tessuto adiposo. Tra i dotti si trova tessuto connettivo lasso, ricco di cellule e vascolarizzato. Dotti e alveoli risultano costituiti da uno strato di cellule epiteliali cubiche o colonnari, rivestito da uno strato di cellule mioepiteliali appiattite: questo strato nei dotti è discontinuo ed è rappresentato da cellule mioepiteliali stellate. Con l'inizio della gravidanza la mammella raggiunge la sua completa maturazione: grazie all'intervento di estrogeni e progesterone si ha ipertrofia di alveoli, dotti e lobuli e perdita di tessuto adiposo interstiziale. Le cellule epiteliali degli alveoli iniziano a sintetizzare i prodotti di secrezione già dal terzo mese, ma la lattogenesi avviene solo in seguito alle modificazioni indotte dal parto. Nei primi giorni successivi al parto le cellule secernenti producono il colostro, sostanza che contiene carboidrati, proteine, immunoglobuline e vitamine e che ha un ruolo molto importante, sia per favorire.
Scene 5 (15m 30s)
[Audio] l'evacuazione e dare inizio ai primi processi digestivi, sia per sviluppare le difese immunitarie del neonato. Dopo questa fase inizia la produzione di latte maturo, indotta dalla caduta del tasso ematico di estrogeni e progesterone, che scatena la produzione di prolattina da parte dell'adenoipofisi. 3. ARTERIE La parete delle arterie è organizzata in tre tonache concentriche: Tonaca intima: formata dalle cellule endoteliali del rivestimento interno, con asse orientato longitudinalmente; Tonaca media: formata da cellule muscolari lisce orientate trasversalmente; Tonaca avventizia: rappresenta lo strato esterno, formata da fibroblasti e fibre collagene orientati longitudinalmente. Tra la tonaca intima e la tonaca media si trova la membrana elastica interna, mentre tra la tonaca media e la tonaca avventizia si trova la membrana elastica esterna. Arterie elastiche Sono le arterie di grosso calibro (7-30 mm di diametro). Alcuni esempi sono: Arteria aorta Vasi arteriosi maggiori (tronco brachiocefalico, arterie carotidi interna ed esterna, arterie succlavia e iliaca) Tronco polmonare Si distendono in fase sistolica e in fase diastolica hanno un ritorno elastico che mantiene costante il flusso ematico. La tonaca intima ha uno spessore di circa 100 micrometri ed è formata da un sottile strato endoteliale, pavimentoso semplice, che poggia su uno strato di tessuto connettivo lasso formato da collagene, fibre elastiche, raramente fibroblasti, macrofagi e cellule muscolari lisce. La tonaca media ha uno spessore di circa 500 micrometri ed è formata da lamine elastiche fenestrate, collegate da sottili fascetti elastici. Tra le lamine elastiche si trovano cellule muscolari lisce disposte circolarmente (per opporsi alla sovradistensione del vaso durante la sistole), fibre collagene ed elastina in una matrice di proteoglicani. La membrana elastica esterna è spesso discontinua..
Scene 6 (17m 42s)
[Audio] La tonaca avventizia è sottile e formata da tessuto connettivo. Presenta fasci longitudinali di fibre collagene e un reticolo sottile di fibre elastiche. È vascolarizzata da una rete di piccoli vasi che prende il nome di vasa vasorum..
Scene 7 (18m 0s)
[Audio] Arterie muscolari Rappresentano la maggior parte delle arterie e presentano un diametro di 0,1-7 mm. La tonaca intima è simile a quella delle arterie elastiche, ma più sottile. Le cellule endoteliali sono poste longitudinalmente e poggiano su un sottile strato endoteliale. La membrana elastica interna appare come uno strato ondulato e presenta fenestrazioni. La tonaca media è costituita prevalentemente da cellule muscolari lisce ad andamento circolare o elicoidale, separate tra loro da una lamina basale che si interrompe a livello dei punti in cui avviene la conduzione intercellulare. Tra le cellule muscolari si intercalano fibre elastiche e fibre collagene di tipo III. La tonaca avventizia è molto sviluppata, formata da fasci di fibre collagene, fibre elastiche e sostanza fondamentale che contiene dermatansolfato ed eparansolfato. Nella parte più esterna si localizzano i vasa vasorum e le terminazioni delle fibre nervose efferenti amieliniche. 4. VENE Hanno in genere una parete più sottile delle arterie, con una distinzione meno netta tra le tre tonache. Si evidenzia un minore sviluppo del contingente elastico, con una prevalenza di fibre collagene. Il tessuto muscolare non ha un ordinamento compatto e i fascetti di miocellule sono separati da maglie di tessuto connettivo. Vene di tipo recettivo Hanno una parete sottile, di tipo fibroelastico e con poche cellule muscolari disposte in modo circolare. Le tonache non sono nettamente distinguibili. Le ritroviamo nei territori sopradiaframmatici, in cui il sangue defluisce verso il cuore favorito dalla forza di gravità e la pressione nel vaso si mantiene bassa. Vene di tipo propulsivo Hanno una parete spessa, ricca di fibre muscolari ed elastiche. Le tonache sono ben riconoscibili e la media è quella più rappresentata. Nella tonaca interna è presente l'endotelio e sotto di esso si trova una spessa lamina di fibre collagene, contenente anche fibre elastiche e fascetti miocellulari. Nella tonaca media si ha prevalentemente la presenza di cellule muscolari, disposte circolarmente o a spirale. La tonaca esterna è ben sviluppata ed è costituita da fibre collagene e fibre elastiche..
Scene 8 (20m 29s)
[Audio] Le vene propulsive sono tipiche dei territori sottodiaframmatici, in cui il deflusso di sangue è contrastato dalla forza di gravità. Il ruolo delle fibre muscolari è proprio quello di favorire il ritorno del sangue al cuore. 5. CUORE La parete del cuore è costituita da tre strati: Epicardio Riveste tutta la superficie esterna del cuore e rappresenta il foglietto viscerale del pericardio sieroso. A livello del peduncolo vascolare si riflette nel foglietto parietale del pericardio sieroso, che tappezza internamente il pericardio fibroso. Superficialmente è costituito da mesotelio, coperto da un velo liquido e dotato di una sottile lamina propria ricca di fibre elastiche; profondamente è congiunto al miocardio tramite lo strato sottoepicardico. Miocardio Rappresenta lo strato più spesso della parete cardiaca ed è formato dal tessuto muscolare striato. Si distinguono il miocardio comune, che assicura l'attività contrattile del cuore, e il miocardio specifico, che rappresenta il sistema di conduzione che trasmette l'impulso contrattile. Miocardio comune: è costituito da elementi mononucleati di forma cilindrica (lunghezza 80100 micrometri, spessore 10-20 micrometri), con le estremità ramificate che prendono contatto con gli elementi adiacenti tramite i dischi intercalari, formando un vero e proprio sincizio funzionale. I cardiociti si riuniscono in fasci tra i quali si intercalano tralci di tessuto connettivo, ricchi di fibre elastiche a livello atriale. I cardiociti presentano un nucleo centrale e numerosi mitocondri e granuli di glicogeno. Le proteine contrattili sono organizzate con la caratteristica striatura trasversale, raccogliendosi in fasci più estesi rispetto alle miofibrille del tessuto.
Scene 9 (22m 22s)
[Audio] muscolare scheletrico. I dischi intercalari sono complessi giunzionali tra le estremità di cardiociti contigui e hanno un andamento scalariforme: i tratti trasversali presentano desmosomi e fasce aderenti, mentre i tratti longitudinali presentano le gap junctions necessarie alla diffusione dello stimolo eccitatorio. I cardiociti atriali (mioendocrini) si distinguono per la presenza di granuli circondati da membrana, contenenti il fattore natriuretico atriale (ANF), che stimola diuresi ed eliminazione di sodio ed è secreto in seguito all'aumento della pressione endoatriale che causa uno stiramento dei cardiociti. Miocardio specifico: costituisce il sistema di conduzione del cuore ed è formato da 3 citotipi o Cellule nodali: situate nei nodi senoatriale ed atrioventricolare, sono elementi fusiformi di dimensioni inferiori rispetto ai cardiociti. Presentano scarsi fasci di miofilamenti, numerosi mitocondri e accumuli di glicogeno. I dischi intercalari sono ricchi di gap junctions. o Cellule di transizione: situate alla periferia dei nodi, hanno caratteristiche strutturali intermedie tra miocardio specifico e comune. o Cellule del Purkinje: costituiscono le reti sottoendocardiche nelle pareti ventricolari e si distinguono dai cardiociti perché sono più grandi e hanno un aspetto pallido, dovuto ai granuli di glicogeno. I miofilamenti sono scarsi e confinati alla periferia delle cellule. Endocardio È una membrana biancastra e liscia che riveste le cavità cardiache. A livello degli orifizi forma pieghe caratteristiche rappresentate dai lembi valvolari. Ha uno spessore che varia da 20 a 500 micrometri, massimo nell'atrio sinistro. È formato da cellule endoteliali poligonali che poggiano su una lamina propria ricca di fibre elastiche e fascetti di miocellule, ma priva di vasi sanguigni. La lamina propria continua nello strato sottoendocardico, costituito da tessuto connettivo lasso. 6. TIMO Il timo è un organo linfoepiteliale situato nel mediastino anteriore. Appare particolarmente sviluppato nei primi anni della vita postnatale, mentre dopo la pubertà va incontro a involuzione trasformandosi in un corpo adiposo retrosternale. È un organo linfoide primario che ha la funzione di promuovere la crescita dei progenitori linfoidi T e il loro differenziamento nei vari tipi di linfociti T (helper, citotossici e regolatori). Ha la forma di una piramide quadrangolare, il cui apice è diviso nei due corni cervicali del timo. Il timo è riccamente vascolarizzato. Anatomia microscopica Il timo è costituito da due lobi, suddivisi in vari lobuli da setti che si dipartono dalla capsula connettivale che circonda l'organo, ricca di fibre elastiche. Ciascun lobulo può inoltre essere diviso in due zone: Regione corticale: regione periferica, molto tingibile e a elevata densità cellulare. È caratterizzata dalla presenza dei linfoblasti, precursori dei linfociti T che giungono al timo tramite i vasi che decorrono nella capsula e nei setti connettivali. Presenta 3 citotipi: o Timociti, accolti tra le maglie del reticolo delle TEC. Vengono in gran parte eliminati per fagocitosi dai macrofagi; o Macrofagi, situati nei pressi dei vasi sanguigni; o Cellule epiteliali timiche (TEC), difficilmente identificabili con le comuni colorazioni ematossilinaeosina. A livello della corticale si portano ad abbracciare i capillari sanguigni, costituendo la barriera.
Scene 10 (26m 10s)
[Audio] ematotimica che protegge i timociti dagli antigeni circostanti. Hanno forma stellata e organizzano una rete tridimensionale, mantenendosi in contatto tramite desmosomi. Regione midollare: regione centrale, più chiara, a densità cellulare minore e non sepimentata dai setti connettivali. È caratterizzata da timociti che migrano dalla corticale e contraggono rapporti più estesi con le cellule epiteliali timiche rispetto alla corticale. Sono presenti anche cellule dendritiche, che possono essere considerate cellule presentanti l'antigene. L'elemento fondamentale per il riconoscimento della regione midollare è rappresentato dai corpuscoli di Hassall: sono formazioni rotondeggianti formate da cellule epiteliali andate incontro a fenomeni di cheratinizzazione, organizzate intorno a una massa granulosa formata da detriti cellulari. Si fanno più numerosi e voluminosi durante l'involuzione dell'organo ed Figura 1: Corpuscolo di Hassall esprimono attivamente citochine e fattori di crescita. 7. MILZA La milza è l'organo linfoide secondario più grande del corpo umano. È situata nell'ipocondrio sinistro, è rivestita dal peritoneo ed è riccamente vascolarizzata: riceve sangue dal tronco celiaco attraverso l'arteria splenica ed è drenata nel circolo portale attraverso la vena splenica. La milza svolge importanti funzioni: provvede alla distruzione degli eritrociti e delle piastrine invecchiate (emocateresi) e ripara gli eritrociti giovani (pitting); agisce da organo linfoide secondario: interviene contro antigeni circolanti nel sangue; funge da serbatoio per piastrine, granulociti ed eritrociti. Ha un colore rosso scuro e una consistenza molle, infatti va spesso incontro a rottura in seguito a traumi addominali. Anatomia microscopica La struttura della milza riflette la sua duplice funzione: la capacità di fornire risposte immunitarie contro gli antigeni circolanti riguarda la polpa bianca, mentre la sua funzione di filtro del sangue spetta alla polpa rossa. La milza è rivestita, sotto il peritoneo, da una capsula connettivale in cui sono accolti fascetti di fibrocellule muscolari lisce e fibre elastiche. Dalla capsula si dipartono corti setti connettivali, detti trabecole, che non dividono il parenchima in lobi e lobuli. All'interno delle trabecole decorrono le arterie trabecolari, che danno luogo ai vasi che forniscono la porzione linfoide dell'organo. Polpa bianca È formata da linfociti e macrofagi, accolti in un fine reticolo connettivale. Si organizza intorno alle arterie follicolari (o centrali della polpa), vasi che derivano dalle arterie trabecolari. Il tessuto linfoide può espandersi costituendo dei follicoli linfatici, denominati corpuscoli lienali del Malpighi: rappresentano l'area B-dipendente della polpa bianca e possono essere primari o secondari. Follicoli primari: accumuli linfoidi rotondeggianti e fortemente basofili (basofilia data dal nucleo voluminoso dei linfociti);.
Scene 11 (29m 27s)
[Audio] Follicoli secondari: presentano una zona mantellare periferica, caratterizzata da piccoli linfociti B, e una zona centrale più chiara denominata centro germinativo, formata da linfociti B che si stanno differenziando in plasmacellule. Essendo le plasmacellule più abbondanti in citoplasma rispetto ai linfociti B, sono meno tingibili e conferiscono al centro germinativo un aspetto più pallido. Figura 2: Follicolo secondario Figura 3: PALS Figura 4: Cordoni di Billroth La polpa bianca si organizza anche in altro modo, costituendo le guaine linfoidi periarteriolari o PALS: rappresentano l'area T-dipendente della polpa bianca. Qui i linfociti T si dispongono a formare dei manicotti linfoidi. Polpa rossa È costituita da una porzione parenchimale e una porzione vascolare, che conferiscono a questa parte dell'organo un aspetto spugnoso. La componente parenchimale è data dai cordoni di Billroth, formati da un reticolo di fibre collagene in cui sono presenti linfociti, macrofagi, ma anche cellule provenienti dal torrente circolatorio come gli eritrociti. La presenza dei macrofagi è legata alla funzione di emocateresi svolta dalla polpa rossa. La componente vascolare invece è costituita dai seni venosi splenici. I primi vasi che forniscono la polpa rossa sono le arterie penicillari, a cui fanno seguito i capillari a guscio: sono capillari privi di rivestimento endoteliale e sono forniti di un involucro di macrofagi disposti intorno al lume del vaso. Sono anche dotati di fibrocellule muscolari lisce, in grado di modulare l'immissione di sangue nella polpa rossa. I capillari a guscio si aprono nei seni venosi splenici, una rete interconnessa di sinusoidi dotata di un endotelio "a doga di botte", formato da cellule allungate e distanziate tra loro con una membrana basale discontinua. Questo endotelio è tale da permettere un transito massivo di elementi figurati, che raggiungono i cordoni di Billroth. I seni venosi splenici si aprono nelle venule della polpa, che a loro volta si aprono nelle vene trabecolari: queste trasportano il sangue venoso refluo verso la vena splenica. 8. LINFONODI I linfonodi sono organi parenchimatosi provvisti di capsula, di forma ovoidale e dimensioni variabili da pochi mm a più di 1 cm. Si trovano sul decorso del sistema vascolare linfatico e hanno il compito di favorire l'incontro tra antigeni trasportati dal circolo linfatico, montare risposte immunitarie e permettere la maturazione dei linfociti B in plasmacellule e cellule della memoria..
Scene 12 (32m 13s)
[Audio] Anatomia microscopica Sono dotati di una capsula di tessuto connettivo denso, arricchita da fibre elastiche e fibrocellule muscolari lisce, che si ispessisce a livello dell'ilo dell'organo. Dalla capsula si dipartono esili setti trabecolari, difficilmente individuabili a causa della popolazione linfoide fittamente stipata. La capsula è attraversata da vasi linfatici in ingresso che convogliano la linfa nel linfonodo: in tal modo gli antigeni presenti in essa interagiscono con la popolazione linfoide B e T dell'organo, andando a determinare espansione clonale e maturazione che permetteranno lo sviluppo di una risposta immunitaria adeguata. Il parenchima è sostenuto da una trama di fibre reticolari (su cui troviamo macrofagi) che supporta e sostiene i linfociti. Si distinguono tre aree: zona corticale, paracorticale e midollare, che costituiscono tre domini funzionali formati da diversi tipi di linfociti. Zona corticale È organizzata in strutture sferoidali ad elevata cellularità (colorazione scura). Questi aggregati di cellule linfoidi prendono il nome di follicoli linfatici e costituiscono le aree B-dipendenti del linfonodo: si distinguono in follicoli primari (inattivi) e secondari (attivi). Questi ultimi sono provvisti di una zona mantellare intensamente colorata e un centro germinativo chiaro. Zona paracorticale È una zona priva di follicoli linfatici: manca di un'organizzazione caratteristica e per questo ha un aspetto più omogeneo. Rappresenta l'area T-dipendente del linfonodo. Zona midollare Questa zona appare chiara, formata da una popolazione linfocitaria variegata che si organizza in cordoni che si ramificano e si interconnettono, sostenuti da fibre reticolari originanti dalla capsula. A livello midollare troviamo le plasmacellule, deputate alla sintesi di immunoglobuline..
Scene 13 (34m 13s)
[Audio] La circolazione linfatica del linfonodo ha origine da numerosi vasi linfatici in ingresso che perforano la capsula e riversano la linfa in una lacuna detta seno sottocapsulare. Da questa lacuna si dipartono canali linfatici intraparenchimali, che invadono dapprima la corticale (seni linfatici corticali) e poi la midollare (seni linfatici midollari). La circolazione intraparenchimale termina a livello dell'ilo, in cui è presente il seno terminale, da cui la linfa abbandona il linfonodo tramite i vasi linfatici efferenti. La circolazione sanguigna del linfonodo è rappresentata da vasi sanguigni che fanno il loro ingresso dall'ilo, formando reti microvascolari capillari da cui originano le venule post-capillari, attraversate da linfociti che entrano nel torrente circolatorio. 9. MALT Il tessuto linfoide associato alle mucose è caratterizzato da diversi livello di organizzazione. È situato nella lamina propria delle tonache mucose, contrae rapporti con l'epitelio e può raggiungere la tonaca sottomucosa. Risulta particolarmente abbondante nel canale alimentare (GALT) e nelle vie respiratorie (BALT). Il ruolo del MALT è quello di garantire l'immunità a livello delle tonache mucose, mediando risposte umorali e cellulari in base alle stimolazioni antigeniche locali. Le risposte umorali si basano principalmente sulla produzione di IgA, mentre le risposte cellulari sono di tipo citotossico. Anatomia microscopica Il MALT può organizzarsi come cellule linfoidi disperse, noduli linfoidi solitari oppure veri e propri organi, come la tonsilla palatina. Si concentra a livello di alcune regioni, come l'anello linfoide della faringe, costituito dalle tonsille palatine, tubariche e linguali, e l'appendice vermiforme. Ai follicoli linfoidi del MALT risulta associato un epitelio particolare (FAE, Follicle-Associated Epithelium) contraddistinto dalla presenza di particolari cellule: Linfociti intraepiteliali Cellule M, formate da una superficie luminale con microvilli e una porzione basolaterale invaginata che accoglie linfociti, macrofagi o APC. Il loro ruolo è quello di prelevare antigeni dal lume e trasferirli nelle tasche citoplasmatiche per favorirne il riconoscimento da parte dei linfociti. Cellule dendritiche, che inviano i prolungamenti tra le cellule epiteliali per prelevare antigeni dal lume e presentarli ai linfociti nella lamina propria..
Scene 14 (36m 55s)
[Audio] Tonsilla palatina La tonsilla palatina è un organo linfoide secondario parzialmente capsulato contenuto tra i due archi glossopalatino e faringopalatino, nell'istmo delle fauci. Fa parte dell'anello orolinfatico di Waldeyer e presenta due regioni: Faccia mediale: formata da epitelio pavimentoso pluristratificato, che presenta profonde invaginazioni a fondo cieco dette cripte tonsillari. Sul fondo delle cripte si organizza un materiale formato da detriti, batteri e linfociti. La porzione superficiale della lamina propria sottostante all'epitelio è occupata da linfociti che in parte migrano verso l'epitelio, mentre la porzione profonda è organizzata in follicoli linfatici di tipo secondario, quindi con la presenza di centri germinativi. Faccia laterale: caratterizzata da una spessa capsula connettivale adagiata su fascetti di muscolatura striata. Dal connettivo capsulare si dipartono setti connettivali. 10. TESSUTO OSSEO Il tessuto osseo è una forma specializzata di tessuto connettivo caratterizzata da una matrice extracellulare mineralizzata, che conferisce all'osso consistenza e resistenza. È formato da più tipi cellulari inseriti in una sostanza intercellulare formata da: Parte organica: formata da collagene di tipo I e sostanza amorfa Parte inorganica: formata da carbonato e fosfato di calcio Anatomia microscopica È possibile distinguere due tipi di tessuto osseo: non lamellare (presente nella vita fetale) e lamellare (presente nell'adulto). Ciascuna lamella è formata da sottili strati di matrice, in cui gli osteociti si dispongono regolarmente e le fibre hanno andamento parallelo. Le lamelle si organizzano in tre tipi di strutture: Osteoni o sistemi di Havers: abbiamo lamelle disposte concentricamente intorno a un canale di Havers, attraversati da vasi e strutture nervose. Altri canali sono i canali di Volkmann, che collegano gli osteoni tra di loro, con il canale midollare e con il periostio. Ogni osteone è separato dai sistemi interstiziali adiacenti tramite la propria linea cementante. Sistemi circonferenziali: lamelle organizzate circolarmente a contatto con le superfici interna ed esterna dell'osso (endostio e periostio). Sistemi interstiziali: lamelle che occupano gli spazi tra gli osteoni. Il tessuto osseo lamellare può a sua volta essere compatto o spugnoso: Tessuto osseo compatto: costituisce la corticale di diafisi ed epifisi nelle ossa lunghe e i tavolati nelle ossa piatte. L'organizzazione è quella spiegata sopra..
Scene 15 (39m 45s)
[Audio] Tessuto osseo spugnoso: costituisce la midollare delle epifisi nelle ossa lunghe. Ha un aspetto alveolare ed è formato da lamelle incomplete che si anastomizzano formando una rete tridimensionale in cui è accolto il midollo osseo. Non vi sono canali in quanto le lamelle sono a contatto con il midollo osseo e vengono nutrite per diffusione. Le cellule del tessuto osseo sono: Osteoblasti: responsabili della produzione della matrice extracellulare e regolano la mineralizzazione dell'osso. Si riscontrano sulle superfici dell'osso in espansione e la loro attività è regolata dall'ormone calcitonina, secreto dalle cellule C della tiroide. Osteociti: elementi quiescenti accolti nelle lacune ossee, forniti di prolungamenti citoplasmatici situati nei canalicoli ossei. Osteoclasti: cellule accolte in fossette scavate sulla superficie della trabecola, che intervengono nel rimodellamento dell'osso con la loro attività fagocitaria. La loro attività è regolata dall'ormone paratiroideo (paratormone)..