Scene 1 (0s)
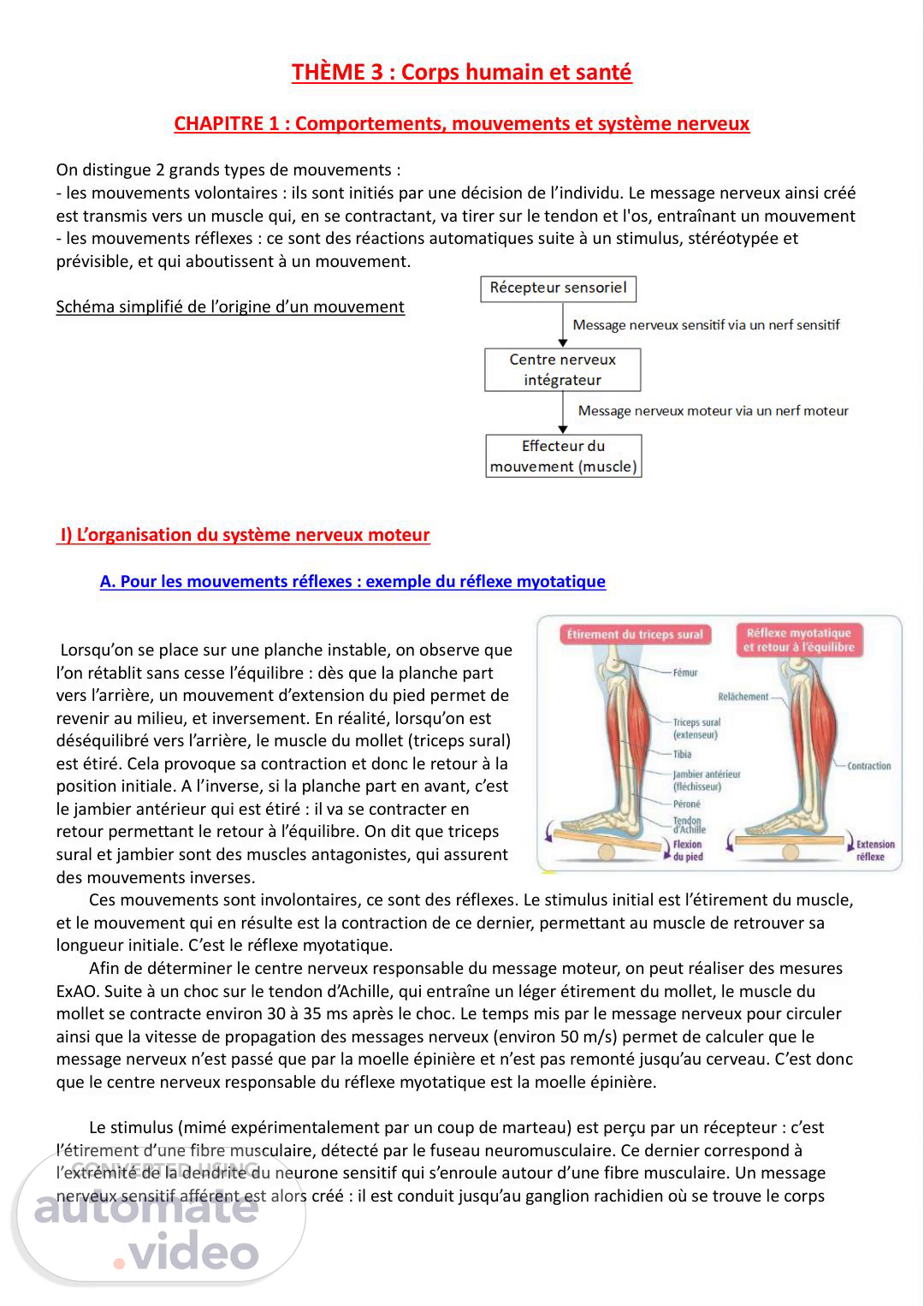
THÈME 3 : Corps humain et santé CHAPITRE 1 : Comportements, mouvements et système nerveux On distingue 2 grands types de mouvements : - les mouvements volontaires : ils sont initiés par une décision de l’individu. Le message nerveux ainsi créé est transmis vers un muscle qui, en se contractant, va tirer sur le tendon et l'os, entraînant un mouvement - les mouvements réflexes : ce sont des réactions automatiques suite à un stimulus, stéréotypée et prévisible, et qui aboutissent à un mouvement. Schéma simplifié de l’origine d’un mouvement I) L’organisation du système nerveux moteur A. Pour les mouvements réflexes : exemple du réflexe myotatique Lorsqu’on se place sur une planche instable, on observe que l’on rétablit sans cesse l’équilibre : dès que la planche part vers l’arrière, un mouvement d’extension du pied permet de revenir au milieu, et inversement. En réalité, lorsqu’on est déséquilibré vers l’arrière, le muscle du mollet (triceps sural) est étiré. Cela provoque sa contraction et donc le retour à la position initiale. A l’inverse, si la planche part en avant, c’est le jambier antérieur qui est étiré : il va se contracter en retour permettant le retour à l’équilibre. On dit que triceps sural et jambier sont des muscles antagonistes, qui assurent des mouvements inverses. Ces mouvements sont involontaires, ce sont des réflexes. Le stimulus initial est l’étirement du muscle, et le mouvement qui en résulte est la contraction de ce dernier, permettant au muscle de retrouver sa longueur initiale. C’est le réflexe myotatique. Afin de déterminer le centre nerveux responsable du message moteur, on peut réaliser des mesures ExAO. Suite à un choc sur le tendon d’Achille, qui entraîne un léger étirement du mollet, le muscle du mollet se contracte environ 30 à 35 ms après le choc. Le temps mis par le message nerveux pour circuler ainsi que la vitesse de propagation des messages nerveux (environ 50 m/s) permet de calculer que le message nerveux n’est passé que par la moelle épinière et n’est pas remonté jusqu’au cerveau. C’est donc que le centre nerveux responsable du réflexe myotatique est la moelle épinière. Le stimulus (mimé expérimentalement par un coup de marteau) est perçu par un récepteur : c’est l’étirement d’une fibre musculaire, détecté par le fuseau neuromusculaire. Ce dernier correspond à l’extrémité de la dendrite du neurone sensitif qui s’enroule autour d’une fibre musculaire. Un message nerveux sensitif afférent est alors créé : il est conduit jusqu’au ganglion rachidien où se trouve le corps.
Scene 2 (1m 5s)
cellulaire du neurone sensitif, puis jusqu’à la moelle épinière en passant par la racine dorsale (ou postérieure) du nerf rachidien, via l’axone du neurone sensitif. Dans la substance grise de la moelle épinière, le neurone sensitif transmet le message à un neurone moteur par l’intermédiaire d’une synapse, au niveau du corps cellulaire. Le message moteur efférent sera ensuite conduit par l’axone du motoneurone jusqu’au muscle en passant par la racine ventrale (ou antérieure) du nerf rachidien. Le muscle reçoit le message nerveux venant du neurone moteur au niveau de la plaque motrice et se contracte en réponse à son propre étirement. Le réflexe myotatique est donc un réflexe monosynaptique (une seule synapse sur le trajet), dont le centre nerveux est la moelle épinière. B. Pour les mouvements volontaires Le centre réflexe du mouvement volontaire est le cerveau. C’est un organe complexe constitué de deux types de cellules, les neurones et les cellules gliales, que l’on peut distinguer lorsqu’on fait un frottis de cerveau coloré au bleu de méthylène. Les neurones sont responsables du traitement et de la propagation des messages nerveux. Le cerveau contient 100 milliards de neurones dont les corps cellulaires se concentrent majoritairement dans la substance grise du cortex cérébral, situé à la périphérie du cerveau. Les cellules gliales sont environ 4 fois plus nombreuses encore que les neurones . On en distingue 3 types : les astrocytes qui interviennent dans la protection, la nutrition et l’activité des neurones ; les oligodendrocytes qui produisent la gaine de myéline autour des axones des neurones, ce qui accélère la conduction du message nerveux ; et les cellules de la microglie qui assurent les défenses immunitaires (phagocytose, CPA). L’identification des zones responsables de la motricité volontaire nécessite la réalisation d’IRM (Imagerie par Résonance Magnétique) et d’IRM fonctionnelles, qui détectent des variations du débit sanguin : toute augmentation du débit sanguin est le signe d’une augmentation de l’activité cérébrale. Les messages nerveux à l’origine des mouvements volontaires partent du cerveau au niveau de l’aire motrice primaire (ou cortex moteur primaire). L’aire motrice primaire de l’hémisphère cérébral droit dirige les mouvements du côté gauche du corps et l’aire motrice primaire de l’hémisphère cérébral gauche commande les mouvements du côté droit du corps. On dit que la commande est controlatérale. Les axones des neurones du cortex moteur, appelés neurones pyramidaux, doivent donc traverser le plan de symétrie durant leur descente, et réalisent ensuite une synapse avec les motoneurones du côté opposé de la moelle épinière. Remarque : Chaque neurone pyramidal commande un ensemble restreint de fibres musculaires, situé dans un muscle donné, par le biais de différents motoneurones. On peut alors dresser la carte motrice du cerveau en cherchant l'emplacement et la taille des zones dédiées à chaque organe. On peut ensuite à partir de cette carte dessiner un homonculus qui garde les proportions de l'aire motrice dédiée. Plus le muscle concerné permet de réaliser des mouvements fins et précis, plus la zone dédiée dans le cerveau sera grande. La carte motrice L’homonculus moteur.
Scene 3 (2m 10s)
II) Le fonctionnement du système nerveux A. La nature du message nerveux Une fibre nerveuse (dendrite ou axone) est chargée négativement par rapport au milieu extérieur : la différence de charge qui existe est appelée potentiel de membrane. Sans aucune stimulation, le potentiel de membrane est de -60 à -70 mV : on parle de potentiel de repos. Si le neurone reçoit une stimulation suffisante, la membrane de la fibre nerveuse sera dépolarisée au- delà d’un certain seuil appelé seuil de dépolarisation. On observe alors une inversion brève (1 à 2 ms) et locale du potentiel membranaire : c’est le potentiel d’action (PA). La propagation de ce PA le long de la fibre nerveuse se fait à l’identique, sans atténuation. Si le neurone n’est pas suffisamment stimulé, la dépolarisation ne permettra pas le déclenchement du PA et disparaîtra rapidement. Au delà de la valeur seuil de stimulation, la réponse de la fibre nerveuse sera toujours la même, sous la forme d’un PA : on dit que les fibres nerveuses répondent à la loi du tout ou rien. L'intensité du stimulus sera alors codée en fréquence de potentiels d'action : plus la stimulation est forte, plus le nombre de PA émis par unité de temps sera importante. La succession de PA qui circule alors le long de la fibre nerveuse est appelée trains de PA. Par exemple, dans le cas du réflexe myotatique, plus on étire sur le muscle, plus la fréquence de PA émis sera importante au niveau de la dendrite du neurone sensitif. Enregistrement du potentiel de membrane du neurone sensitif lors de stimulations d’intensité croissante Rm : dans les résultats d’expérience, les PA sont souvent figurés par un simple trait Lorsque le message nerveux parvient au niveau d’une synapse, sa propagation électrique devient impossible : il va changer de nature. L’extrémité de l’axone du neurone pré-synaptique contient des vésicules, renfermant des molécules de neurotransmetteurs (= neuromédiateurs). L’arrivée d’un message nerveux électrique provoque la migration des vésicules dans le milieu intérieur du neurone jusqu’à la membrane présynaptique avec laquelle elles fusionnent. Les NT se trouvant dans ces vésicules sont alors déversés dans la fente synaptique : c’est l’exocytose. Au niveau d’une synapse, le message nerveux électrique présynaptique codé en fréquence de PA est traduit en message chimique codé en concentration de NT : plus l’intensité de la stimulation est forte, plus la fréquence des PA présynaptiques est importante, plus la concentration en NT dans la fente synaptique est importante. Suite à l’exocytose, les NT traversent la fente synaptique et se fixent sur les récepteurs spécifiques, présents sur la membrane postsynaptique. Cette fixation provoque une modification de l’état électrique du Un PA.
Scene 4 (3m 15s)
neurone post-synaptique (le motoneurone dans le cas du réflexe myotatique comme d’un mouvement volontaire). Chaque neurone ne libère qu’un seul type de NT. Certains entraînent une dépolarisation de l’élément post-synaptique : ils sont excitateurs, comme l’acétylcholine ou le glutamate. D’autres provoquent une hyperpolarisation de la membrane post-synaptique : ils empêchent l’excitation du neurone et donc dits inhibiteurs, comme le GABA. Les NT se fixent au récepteur postsynaptique de façon brève : ils se décrochent rapidement et se retrouvent ensuite libre dans la fente synaptique où ils seront détruits ou recapturés. Cela permet l’arrêt de la transmission du message nerveux au niveau synaptique. B. Le rôle intégrateur du motoneurone Le corps cellulaire du motoneurone reçoit des informations diverses provenant de synapses excitatrices ou inhibitrices. Ces informations peuvent arriver en même temps, auquel cas il réalise une sommation spatiale, ou bien arriver les unes après les autres, auquel cas il assure une sommation temporelle. Si la dépolarisation de la membrane du corps cellulaire du motoneurone est suffisante, suite à ces sommations, alors il émettra un message nerveux moteur unique, qui correspondra à l’intégration de l’ensemble des informations reçues : on parle d’intégration nerveuse. Ainsi, un motoneurone peut recevoir des messages à la fois excitateurs au niveau de la synapse avec un neurone pyramidal et des messages inhibiteurs au niveau de la synapse avec un interneurone. En fonction de l’intensité du message émis au niveau de ces 2 synapses, le muscle pourra soit se contracter, soit rester relâché. Un exemple concret : Ce qu’il se passe quand on fléchit volontairement le pied : Le message issu des neurones pyramidaux stimule le motoneurone du jambier via une synapse excitatrice, permettant alors la contraction du jambier. En même temps, il faut relâcher le soléaire (mollet) : le neurone pyramidal vient exciter un interneurone qui inhibe le motoneurone du soléaire. Ce qu’il se passe quand on provoque le réflexe myotatique avec le marteau : Le message issu du neurone sensitif stimule le motoneurone du soléaire via une synapse excitatrice , permettant la contraction du soléaire. En même temps il faut relâcher le jambier : le neurone sensitif stimule un interneurone qui inhibe le motoneurone du jambier. Quand on réalise à la fois la flexion volontaire et le réflexe, le résultat est tout de même le maintien de la flexion du pied : le message nerveux issu du neurone pyramidal est « plus fort » que le réflexe : - le motoneurone du soléaire qui aurait dû se relâcher suite au réflexe est plus stimulé par le neurone pyramidal qu’inhibé par l’interneurone - le motoneurone du jambier qui aurait dû se contracter est en fait plus inhibé par l’interneurone que stimulé par le neurone sensitif → il ne se contracte pas (ou très peu) → le réflexe est donc inhibé. C. Du message moteur à la contraction musculaire. Une fois le message nerveux émis au niveau du corps cellulaire du motoneurone, il va se propager le long de l’axone et parvenir au niveau de la plaque motrice : c’est une synapse entre l’extrémité axonique du motoneurone et une fibre musculaire..
Scene 5 (4m 20s)
L’arrivée d’un train de PA sur l’extrémité axonique du motoneurone déclenche l’exocytose des vésicules d’acétylcholine au niveau de la fente synaptique. Sa fixation sur les récepteurs de la membrane de la cellule musculaire entraîne une dépolarisation de cette dernière. Si la dépolarisation dépasse un certain seuil de stimulation, un PA musculaire sera créé, se propagera sur toute la membrane et gagnera les tubules T (ou tubules transverses), replis membranaires rapprochant la membrane cellulaire de celle du réticulum sarcoplasmique. Cet organite, baignant dans le cytoplasme, est un compartiment dont la concentration en ions calcium est supérieure à celle du cytoplasme et qui possède des canaux calciques permettant le passage de ces ions. Lors de l’arrivée des PA musculaires à proximité de la membrane du réticulum sarcoplasmique, les canaux calciques s’ouvrent et le calcium est libéré dans le cytoplasme. C’est cet afflux d’ions Ca2+ qui provoque la contraction de la cellule musculaire. Petit point culture : le curare. Le curare est un produit naturel, issu des lianes grimpantes que l'on trouve au cœur des forêts d'Amazonie, et plus particulièrement l'espèce « Chondrodendron tomentosum ». Les guerriers s'en servaient pour concevoir des flèches empoisonnées, capables de paralyser les ennemis touchés. Le curare est en réalité capable de se fixer sur les récepteurs à Ach sans provoquer le dépolarisation : il prend la place de l’Ach et bloque donc le fonctionnement de la plaque motrice sur une durée prolongée. Il empêche progressivement tous les muscles de fonctionner, dont le diaphragme (délai : environ 2 à 3 mn) : il entraîne alors la mort par asphyxie. Aujourd’hui, on utilise des analogues du curare en médecine pour les anesthésies, mais son utilisation est forcément couplée à une intubation puisque la respiration naturelle n’est plus possible !! L’intégrité du système nerveux doit être maintenue si on veut que le fonctionnement soit optimal. Par exemple, dans certains cas d’accidents, la moelle épinière est lésée voire rompue : les mouvements réflexes sont encore possibles (puisque le message ne remonte pas jusqu’au cerveau), mais la motricité volontaire en-dessous de la lésion devient impossible puisque le message nerveux ne peut plus circuler. En fonction de la localisation de la lésion, la paralysie est plus ou moins étendue..
Scene 6 (5m 25s)
En plus des accidents, l’âge et la consommation de certaines substances peuvent altérer le fonctionnement du système nerveux, et notamment du cerveau de façon plus ou moins prolongée… III) Le cerveau, un organe fragile à préserver Le cerveau possède une capacité à se réorganiser après un traumatisme (AVC…) ou après une opération chirurgicale : cette capacité est appelée plasticité cérébrale. Les mécanismes à l’œuvre sont de mieux en mieux connus, et on observe : - des transferts de compétences de zones corticales (comme dans le cas d’Arthur, enfant épileptique) : la zone lésée ne pouvant plus assurer sa fonction, c’est une autre zone du cerveau qui, après plusieurs mois, pourra contrôler cette fonction, mais de façon souvent imparfaite, et sans perdre sa fonction initiale - la formation de nouvelles synapses entre des neurones qui n’étaient pas connectés au départ ou l’activation de réseaux de neurones existants mais non fonctionnels auparavant, comme c’est le cas suite à des entraînements. Par exemple, 3 zones ont montré une augmentation de volume chez les musiciens par rapport à des novices : aires motrices, aires auditives et aires associatives. Le temps de réponse et l’efficacité des voies motrices sont améliorés chez les musiciens : renforcement de l’efficacité et du nombre de synapses pour une même zone de corps (par exemple l’index). L’IRM ci-contre révèle une fracture sévère de la 7ème vertèbre cervicale avec atteinte de la moelle épinière (en rouge). Les lésions de ce segment particulièrement vulnérable de la colonne vertébrale (accidents de la circulation, chutes) sont la cause de paralysies graves ou de décès. L’homme ici a perdu l’usage de ses jambes et de ses bras, il est tétraplégique..
Scene 7 (6m 29s)
Mise en évidence de la plasticité cérébrale chez un pianiste.